
Pilus Assembly in Bacterial Pathogens
You are here
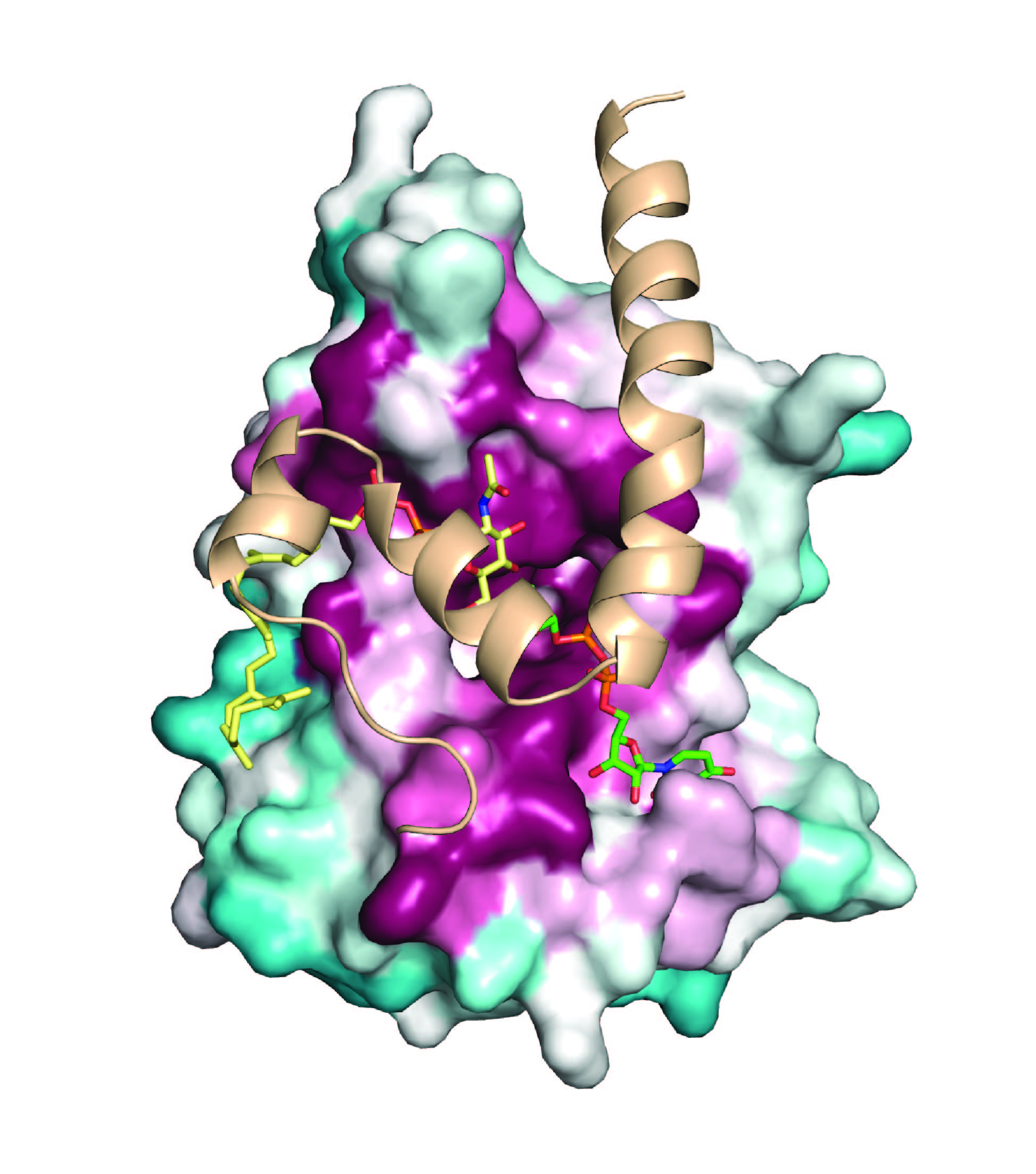
Bacterial pathogens display virulence factors that enable them to adhere to host tissues, resist phagocytic killing, invade host cells and acquire essential nutrients. We are studying how bacteria synthesize and display two types of virulence factors: (i) pili, proteinaceous fibers that project from the bacterial cell surface to mediate adhesion, and (ii) Wall Teichoic Acids (WTAs), highly abundant cell wall anionic glycopolymers that regulate cell division, and other fundamental aspects of bacterial physiology. A greater understanding of these processes could lead to new antibiotics to treat infections caused by multi-drug resistant bacteria.
(i) Pili and protein display- Sortase enzymes are key architects of the Gram-positive bacterial cell wall that assemble pili and display proteins. Our prior research has helped to elucidate how these enzymes covalently attach proteins to the cell wall and how they recognize their substrates. In exciting new studies, we are investigating how Gram-positive bacteria use sortases to construct pili: long, thin fibers (0.2–3.0 µm x 2-6 nm) that project from the cell surface to mediate bacterial adherence to host tissues, invasion, aggregation, biofilm formation, and modulation of host immunity. These protein-structures are distinct from pili produced by Gram-negative bacteria because their protein subunits (called pilins) are crosslinked by lysine-isopeptide bonds that confer enormous tensile strength. Working with the Ton-That group we have recently begun to learn how sortase “pilin polymerases” build pili by covalently linking adjacent pilin subunits together via lysine-isopeptide bonds. Elucidating the molecular basis of this conserved assembly mechanism could have a broad impact, as Gram-positive pili are used by pathogenic microbes to facilitate infections. These pathogens include among others: Corynebacterium diphtheriae, Actinomyces oris, Enterococcus faecium, Streptococci sp. and Bifidobacterium bifidum.
(ii) Wall Teichoic Acid Biosynthesis- One of the most abundant polymers displayed on the surface of Gram-positive bacteria is Wall Teichoic Acid (WTA), an anionic glycopolymer that has critical functions in cell division, morphology, adhesion, and microbial susceptibility to the immune response. The WTA biosynthetic pathway in Staphylococcus aureus has drawn significant interest as a drug target, as clinically important Methicillin Resistant S. aureus (MRSA) strains that lack WTA are defective in host colonization and re-sensitized to b-lactam antibiotics. WTA in S. aureus is comprised of repeated ribitol-phosphate groups that are joined to the peptidoglycan via a conserved disaccharide linkage unit. The polymer is synthesized on the cytoplasmic face of the cell membrane via a multi-step process that elaborates a membrane-anchored undecaprenyl-phosphate carrier lipid. The assembled polymer is then glycosylated, transferred across the cell membrane and covalently attached to the peptidoglycan. Despite its fundamental importance and potential as a drug target, we know surprising little about the enzymatic reactions that construct the conserved linkage unit. In ongoing research, we are therefore using structural, biochemical and cellular approaches to study TagA, a novel glycosyltransferase that catalyzes the first committed step in WTA biosynthesis. The results of these studies will provide important insight into the mechanism of WTA biosynthesis and could facilitate the discovery of new antibiotics that work by inhibiting the biosynthesis of this important polymer.
Relevant publications: